水稻多酚氧化酶基因功能标记的开发与应用_杨杰

水稻多酚氧化酶基因功能标记的开发与应用
杨杰1曹卿2王军1范方军1张玉琼2仲维功1,*
(1江苏省农业科学院粮食作物研究所,江苏南京210014;2安徽农业大学生命科学学院,安徽合肥230036;*通讯联系人,E-mail: wgzhong0503@yah https://www.wendangku.net/doc/081371257.html,)
Development and A pplicat ion of Functional M arkers for Polyphenol Oxidase(PPO)Alleles in Rice
Y ANG Jie1,C AO Qing2,W ANG Jun1,F AN Fang-jun1,Z H ANG Yu-qiong2,Z H ONG We-i gong1,*
(1In stitute of Food C rop s,J iangsu A cad emy of A gr icultur al S ciences,N anj ing210014,C hina;2Colleg e of L if e S cience,A nhui A g ricul-tu ral University,H ef ei230036,China;*Corresp ond ing auth or,E-mail:w g zh ong0503@yah https://www.wendangku.net/doc/081371257.html,)
Y ANG Jie,CAO Qing,WANG Jun,et al.Development and application of functional markers for polyphenol ox idase(PPO) alleles in rice.Chin J Rice Sci,2011,25(1):37-42.
Abstract:T he phenol r eaction(PHR)phenotype is one of the criteria of r ice classification and the gene contro lling PHR has been identified.T he positive PHR phenotype is conditioned by functional polyphenol oxidase(PPO)gene,and negative PHR pheno ty pe is conditioned by non-functional PPO gene.T o seek the markers for rice classification,two InDel markers,
FMppo18and FM ppo29,were developed flanking on the18bp or29bp deletion in ex on3o f PPO alleles.F orty-five rice ma-terials were g enotyped with the two mar kers and their PHR phenotypes were also investigated.Three accessions of w ild r ice and six indica rice varieties car ried non-deletion functional PPO alleles.Among the11japonica rice materials,nine car ried18 bp deletion allele,and two carr ied29bp deletion alleles.H owever,the indica r ice variety Dular carr ied29bp deletion allele and Long qing carried18bp deletion allele.Among the23w eedy r ice accessions,three accessions of Ludao collected from L-i any ung ang of Jiangsu Province,China,and four accessions of T ang dao co llected fr om Huaiyuan,Laian,Q uanjiao,and Fe-i dong of Anhui Pr ovince,China in1950s car ried non-deletion PPO alleles.Among the16weedy r ice accessions collected in re-cent year s,14carried nor mal PPO alleles,and tw o carried18bp deletion PPO allele which was consistent with the cultivated rice in Jiangsu Province.T hese results showed that the positive PHR materials carr ied normal PPO alleles,and the negative PHR mater ials carried deletion PPO alleles,indicating that the phenotype per fectly matched with the genotype.T herefore, the tw o markers can be used in germplasm screening,marker-assisted selection and evo lution research in rice.
Key words:polyphenol ox idase;phenol r eaction;functional marker;w eedy rice;gene flow
杨杰,曹卿,王军,等.水稻多酚氧化酶基因功能标记的开发与应用.中国水稻科学,2011,25(1):37-42.
摘要:水稻酚反应是水稻分类的重要依据。控制水稻酚反应的多酚氧化酶基因(PPO)已经克隆,根据其等位基因的DNA 序列差异设计了InDel功能标记FMppo-18和FMppo-29。利用这两个标记对45份材料进行了标记基因型分析和酚反应鉴定。结果发现3份普通野生稻为非缺失带型;8份籼稻材料中6份为非缺失带型,而Dular为缺失29bp带型,龙晴为缺失18 bp带型,表明Dular和龙晴分别携带29bp和18bp缺失的PPO等位基因;11份粳稻材料中9份携带18bp缺失的PPO等位基因,2份携带29bp缺失的PPO等位基因。对23份杂草稻鉴定的结果表明,7份早年发现的杂草稻,即3份江苏省连云港穞稻和4份安徽省怀远、来安、全椒、肥东的塘稻为非缺失带型;而16份近年来发现的杂草稻中14份不缺失,2份表现为18bp缺失的PPO等位基因,与江苏省大面积推广的栽培粳稻品种携带相同的PPO基因。酚反应结果表明带29bp或18 bp缺失的PPO等位基因的材料酚反应呈阴性,而不缺失的材料酚反应呈阳性。标记基因型与酚反应表现型完全对应。表明这两个功能标记可用于水稻种质资源鉴定、进化研究以及多酚氧化酶基因的分子标记辅助选择育种。
关键词:多酚氧化酶;酚反应;功能标记;杂草稻;基因漂移
中图分类号:Q943.2;S511.032文献标识码:A文章编号:1001-7216(2011)01-0037-06
水稻可分为籼稻和粳稻两个亚种。这两个亚种至少在40多个形态、生理、生态性状上存在差异[1-2],通常以稃毛长、KClO3抗性、酚反应和耐冷性这4个性状来区分籼稻和粳稻,这些性状与水稻驯化密切相关[3-4]。水稻酚反应(phenol r eaction, PH R)是指水稻稻壳在苯酚溶液中的颜色变化,若变为深褐色或黑色称为PH R阳性,若不变色,称为PH R阴性。已有的研究表明,水稻酚反应由一个符合孟德尔定律的单基因(P hr1)控制,位于第4染色体的长臂上[5-8]。Yu等[9]克隆了水稻酚反应基因P hr1,编码多酚氧化酶(po lyphenol oxidase, PPO)。该基因功能正常,酚反应呈阳性;该基因功能缺失,酚反应呈阴性[9-10]。多酚氧化酶基因可能
收稿日期:2010-03-27;修改稿收到日期:2010-05-18。
基金项目:国家转基因生物新品种培育科技重大专项资助项目(2009ZX08001-005B,019);国家科技支撑计划资助项目(2006BADO2A03);江苏省自然科学基金资助项目(BK2006159, 2009321);江苏省政府留学奖学金资助项目;江苏省/六大人才高峰0项目(2010);江苏省农业科学院基金资助项目(6110703, 6510806)。
37
中国水稻科学(Chin J Rice Sci),2011,25(1):37~42 http://www.r https://www.wendangku.net/doc/081371257.html,
DO I:10.3969/j.issn.1001-7216.2011.01.006
与水稻等植物抗病虫以及储藏等特性有关[11-13],因此,开发与筛选基于PCR成本较低的分子标记在育种中具有很大的应用潜力。
功能基因的克隆为开发基因功能标记奠定了基础。基因功能标记(functional m arker)是指一个分子标记位点代表一个特定的等位基因,并与该基因控制的性状相联系,通过对分子标记的筛选即能对性状进行筛选[14-16]。本研究针对多酚氧化酶等位基因的功能基序设计了基于PCR的InDel标记,可用于水稻种质资源的多酚氧化酶基因型鉴定、水稻进化研究以及分子标记辅助选择育种。
1材料与方法
1.1水稻材料
供试材料共计45份,包括3份由广西农业科学院提供的普通野生稻,江苏省农业科学院粮食作物研究所保存的19份品种资源,江苏连云港穞稻3份[17]、安徽怀远塘稻、来安塘稻、全椒塘稻、肥东塘稻各1份,作者近年来在江苏收集到的/红米0杂草稻材料16份(表1)。
1.2酚反应
酚反应参照Oka等[18]的方法。试管中加入3 m L1.5%的苯酚溶液,放饱满种子3~5粒,28e下放置48h,查看颖壳变色程度,变色记为阳性,不变色记为阴性。
1.3InDel标记的引物设计和合成
在GenBank中(http://w w w.ncbi.nlm.nih.
g ov/entr ez/query.fcg i)搜索编码水稻籼稻和粳稻P PO基因的序列。粳稻日本晴的登录号为DQ532376,籼稻9311的登录号为DQ532391,籼稻
表1供试材料
T able1.List o f rice materials us ed in this study.
编号Code
材料
Material
来源
Origin
分类
T axon
备注
Note
1Kasalath印度In dia籼Indica
2Dular印度In dia籼Indica
3IR36国际水稻研究所IRRI籼Indica
4IR24国际水稻研究所IRRI籼Indica
5扬稻6号Yan gdao6江苏扬州Yangzhou,China籼Indica
6南京11Nanjing11江苏南京Nanjin g,China籼Indica
7南京14Nanjing14江苏南京Nanjin g,China籼Indica
8龙晴Longqing江苏南京Nanjin g,China籼Indica
9日本晴Nipponb are日本Jap an粳Japonica
10爱知106Aich i106日本Jap an粳Japonica
11落霜青Luoshuangqing江苏苏州S uzhou,Chin a粳Japonica
12田鸡青T ian jiqing江苏苏州S uzhou,Chin a粳Japonica
13小青种Xiaoqingzh ong江苏苏州S uzhou,Chin a粳Japonica
14白壳老来青Baikelaolaiqing江苏苏州S uzhou,Chin a粳Japonica
15苏御糯Su yunuo江苏苏州S uzhou,Chin a粳Japonica
1602428江苏南京Nanjin g,China粳Japonica
17武运粳7号Wu yunjing7江苏武进Wujin,Chin a粳Japonica
18南粳47Nanjing47江苏南京Nanjin g,China粳Japonica
19武香粳14Wuxian gjing14江苏武进Wujin,Chin a粳Japonica
20-22普通野生稻Common w ild rice广西Guangxi,Chin a普通野生稻Oryz a ru f ip og on3份3accessions 23-25穞稻Ludao江苏连云港Lianyun gang,China杂草稻O.sativa f.sp ontan ea3份3accessions 26怀远塘稻H uaiyuan tangdao安徽怀远H uaiyuan,C hina杂草稻O.sativa f.sp ontan ea
27来安塘稻Laian tangdao安徽来安L aian,C hina杂草稻O.sativa f.sp ontan ea
28全椒塘稻Quanjiaotangdao安徽全椒Qu an jiao,China杂草稻O.sativa f.sp ontan ea
29肥东塘稻Feidon gtangdao安徽肥东Feidong,China杂草稻O.sativa f.sp ontan ea
30-35扬中红米稻Yan gzhong red rice江苏扬中Yangzhon g,Chin a杂草稻O.sativa f.sp ontan ea6份6accessions 36姜堰红米稻Jiangyan red rice江苏姜堰Jian gyan,China杂草稻O.sativa f.sp ontan ea
37如东红米稻Rudong r ed rice江苏如东Rudong,China杂草稻O.sativa f.sp ontan ea
38淮安红米稻H uaian red rice江苏淮安H uaian,Ch ina杂草稻O.sativa f.sp ontan ea
39如皋红米稻Rugao red rice江苏如皋Rugao,Ch ina杂草稻O.sativa f.sp ontan ea
40盐都红米稻Yan du red rice江苏盐都Yandu,China杂草稻O.sativa f.sp ontan ea
41高邮红米稻Gaoyou red rice江苏高邮Gaoyou,Ch ina杂草稻O.sativa f.sp ontan ea
42兴化红米稻Xinghua red rice江苏兴化Xinghu a,China杂草稻O.sativa f.sp ontan ea
43灌云红米稻Guanyun red rice江苏灌云Guanyun,Chin a杂草稻O.sativa f.sp ontan ea
44洪泽红米稻H ongze red rice江苏洪泽H ongze,C hina杂草稻O.sativa f.sp ontan ea
38中国水稻科学(Chin J Rice Sci)第25卷第1期(2011年1月)
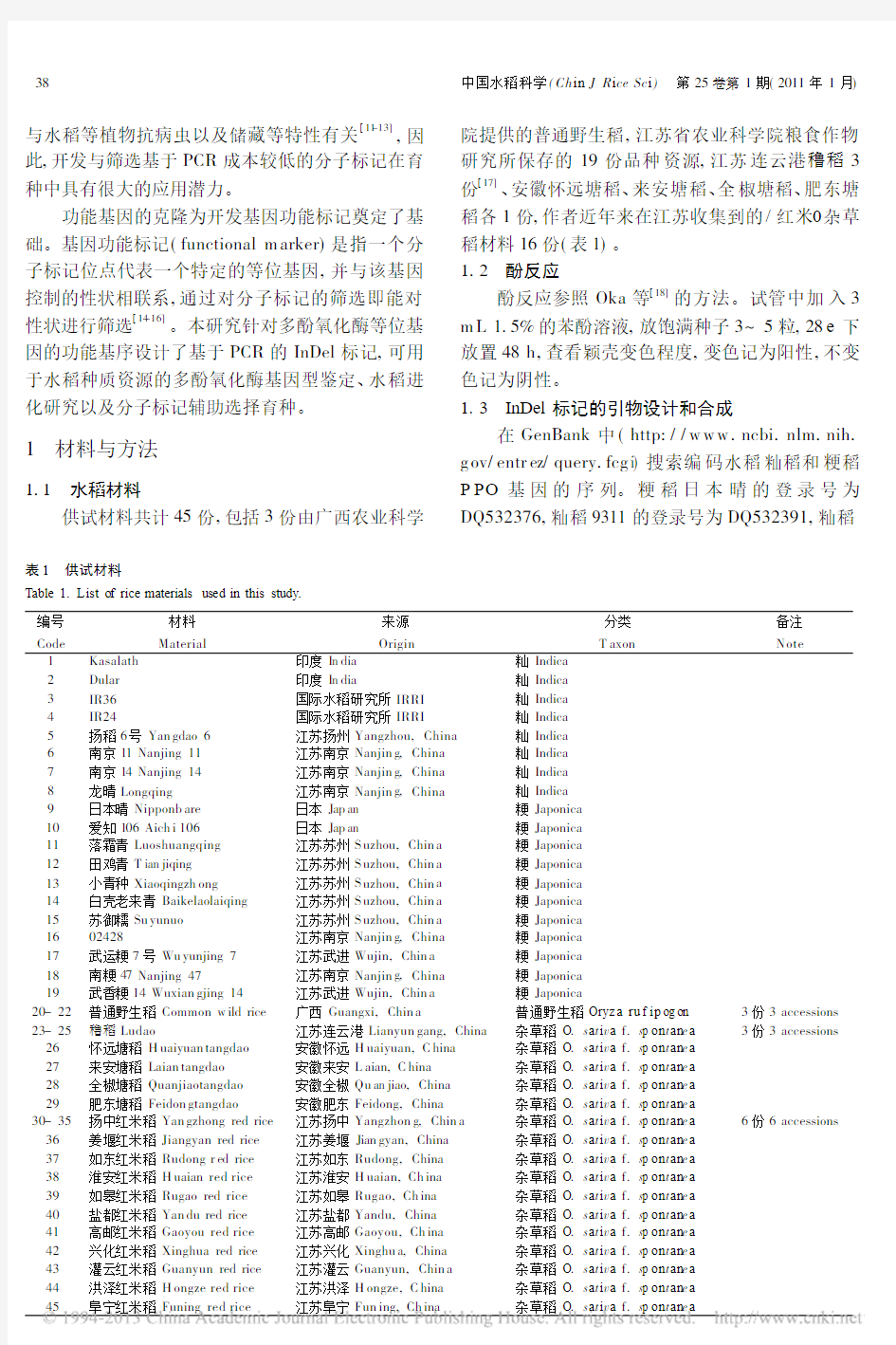
图1 PPO 及其等位基因的结构
Fig. 1.Structur e of the PPO allele locus.
正常的P PO 基因具有3个外显子和2个内含子。功能缺失的P PO 基因主要是第3外显子存在部分碱基的缺失:一类是18bp 缺失;一类是29bp 缺失。明恢63携带正常P PO 基因,02428携带29bp 缺失的等位基因,日本晴携带18bp 缺失的等位基因。
Th e functional P PO gene consists of 3exons (grey boxes)and 2introns (w hite boxes)w ith its start codon (ATG)in exon 1and stop co -don (T GA)in exon 3.Th e null p p o -E3-18allele h as an 18bp deletion in ex on 3,and th e null p po -E3-29allele has a 29bp deletion.T he rice variety Min gh ui 63has the functional PP O allele,the rice variety Nipponb are w ith negative phenol reaction(PHR)h as the nu ll p p o -E3-18a-l lele,and the rice variety 02428with negative PH R has th e null p p o -E3-29allele.
明恢63的登录号为DQ532375,粳稻Sun 的登录号为DQ532383。籼稻没有缺失,粳稻缺失29或18对碱基。为了检测和区分粳稻和籼稻P PO 基因型,选取发生了缺失区段的两侧400~500bp 范围内的序列,用Pr im er Pr em ier 5软件设计引物,使其扩增的PCR 产物大小适中,能较好地在琼脂糖凝胶上电泳分析。引物由Invitrog en 公司合成。1.4 DNA 提取、PCR 扩增、电泳及结果记录利用SDS 法提取总DN A 。以总DNA 为模板,按下列反应体系进行PCR 。PCR 反应体系(20L L)为:DNA 1.0L L,上游引物0.5L L,下游引物0.5L L,10@缓冲液2.0L L,M gCl 2(5m mo l/L)2L L,Taq 酶0.2L L,dNT P (2.5mm ol/L)1L L,ddH 2O 12.8L L 。扩增条件为:94e ,5min;94e ,30s,55e ,30s,72e ,30s,35个循环;72e ,延伸10min 结束反应。PCR 产物在含有溴化乙淀(EB)的3.0%琼脂糖凝胶中120V 下电泳40min,然后在凝胶成像系统中拍照。
2 结果与分析
2.1 InDel 标记的开发
根据Yu 等[9]的研究结果,绝大多数籼稻携带
功能正常的多酚氧化酶基因,而粳稻携带功能缺失的多酚氧化酶基因。粳稻中主要有两个多酚氧化酶等位基因:一个是第3外显子缺失29bp;一个是第3外显子缺失18bp 。根据水稻多酚氧化酶等位基因结构的差异(图1),设计了两个多酚氧化酶等位基因的功能标记引物FM ppo -29和FM ppo -18。引物序列与扩增产物片段大小见表2。
2.2 InDel 标记FMppo -18和FMppo -29对45份水稻材料的基因型检测结果
从图2和图3可见,供试材料中传统分类为籼稻的品种南京11、南京14、扬稻6号、IR24、IR36、Kasalath,FMppo -18和FM ppo -29标记PCR 产物的带型为非缺失带型,3份来自广西的普通野生稻也为非缺失带型,说明这些材料携带正常功能的P PO 基因;FMppo -18标记对传统分类属于粳稻的材料如太湖流域粳稻资源落霜青、田鸡青、小青种、白壳老来青,日本的粳稻品种日本晴、爱知106,江苏的粳稻品种如武运粳7号、武香粳14、南粳45、南粳47,PCR 产物的带型为缺失带型,而02428和苏御糯为非缺失带型,表明多数粳稻携带18bp 缺失的等位基因;FMppo -29标记检测结果表明02428和苏御糯为缺失带型,其他粳稻材料为非缺失带
表2 ppo -E3-29和ppo -E3-18等位基因功能标记开发
T able 2.Development of functional markers for bo th the p po -E 3-29and p p o -E3-18alleles.
PPO 等位基因PPO allele
序列差异
Sequence divergence
功能标记Functional m arker 引物序列
Pri m er sequence (5c -3c )
P CR 产物
PCR product 退火温度T m/e
p p o -E3-29外显子3中29bp 缺失FMppo -29F:
GT GT T CT T CGCGCACCA 115/96bp 66A 29bp deletion in exon 3R:AGAAGAAGCTGGCGTCGA (PPO /p p o -E3-29)p p o -E3-18
外显子3中18bp 缺失FMppo -18
F:GT T CCCGGCGACCCT GGACA
143/125bp 55
An 18bp deletio n in ex on 3
R:
ACTT GACGTA CGTGGAGT GGT CGGG
(PPO /p p o -E3-18)
39
杨杰等:水稻多酚氧化酶基因功能标记的开发与应用
图2 功能标记FMppo -18对45份材料的检测结果
Fig. 2.Pr ofile of the FM ppo -18P CR amplification pr oducts from genomic DN As of 45r ice materials.
M -DNA 分子量标准;图中编号与表1相同。图3同。
M ,DNA ladder m ark ers;Codes are same as in Table 1.T he same as in Fig.
3.
图3 功能标记FMppo -29对45份材料的检测结果
Fig. 3.Pr ofile of the FM ppo -29P CR amplification pr oducts from genomic DN As of 45r ice materials.
型,表明这两个粳稻材料携带29bp 缺失的P PO 等位基因。龙晴、Dular 传统分类为籼稻,用FMppo -18标记对龙晴的检测结果为18bp 缺失带型,而Dular 为29bp 缺失带型。
两个标记对23份杂草稻鉴定结果表明,江淮流域早年发现的杂草稻即3份穞稻、4份塘稻均为非缺失带型,而近年来发现的16份杂草稻资源中,除淮安杂草稻和灌云杂草稻的FM ppo -18标记检测结果为缺失带型外,其他14份杂草稻材料两个标记均为非缺失带型。说明23份杂草稻材料中除2份携带功能缺失的PP O 等位基因外,其他杂草稻都携带功能正常的PP O 基因。有意思的是,这2份近年来在江苏省收集的杂草稻携带的P PO 等位基因与江苏省的栽培品种携带相同的PP O 基因,即都是18bp 缺失的PP O 等位基因。2.3 酚反应结果
45份材料在酚溶液中处理48h 后,太湖流域粳稻资源苏御糯、落霜青、田鸡青、小青种、白壳老来青、02428,日本的粳稻品种日本晴、爱知106,江苏
的粳稻品种武运粳7号、武香粳14、南粳47,淮安红米杂草稻,灌云红米杂草稻,龙晴,Dular,这15份材料的颖壳不变色;5份籼稻、3份普通野生稻、3份穞
稻、4份塘稻、14份红米稻,这29份材料的颖壳都变为棕褐色。45份供试材料的基因型检测结果和酚反应表现型结果见表3。从结果可以看出,酚反应呈阳性的水稻材料都携带功能正常的P PO 等位基因,酚反应呈阴性的水稻材料都携带功能缺失的P PO 等位基因,标记基因型与酚反应表现型完全对应。传统分类为籼稻的IR24,48h 酚反应后,两片护颖和稃尖均能被染色,而稻壳略有变色,继续处理72h 后,稻壳颜色变深(图4),其原因有待进一步研究。
3 讨论
3.1 基因功能标记的开发
Andersen 等[14]
首先提出基因功能标记概念,功能标记的多态性来源于造成等位基因功能差异的DNA 序列差异,可以进行基因型鉴定和基因型选
40中国水稻科学(Chin J Rice Sci) 第25卷第1期(2011年1月)
表345份材料的基因型与表现型
T able3.Genotypes o f the45rice ma terials tested.
等位基因Allele
序列差异S equence divergence
29bp缺失
29bp deletion
18bp缺失
18bp deletion
水稻材料
Rice material
酚反应
Phenol reaction
PP O否No否No Kasalath、IR36、扬稻6号、南京11、南京14、IR24、3份普通野生稻、
3份穞稻、4份塘稻、14份红米杂草稻
Kasalath,IR36,Yangdao6,Nan jing11,Nanjing14,IR24,3ac-
cessions of Guan gxi com mon w ild rice,3accessions of Ludao,4ac-
cessions of T an gdao,14accessions of weedy red rice
阳性Positive
p p o-E3-29是Yes否No02428、苏御糯、Dular
02428,S uyunuo,Dular
阴性Negative
p p o-E3-18否No是Yes日本晴、爱知106、武运粳7号、武香粳14、南粳45、南粳47、落霜青、
田鸡青、小青种、白壳老来青、龙晴、淮安杂草稻和灌云杂草稻
Nipponbare,Aichi106,Wu yunjing7,Wuxiangjing14,Nanjing
45,Nanjing47,Luoshuangqing,T ian jiq ing,Xiaoqin gzhong,
Baik elaolaiq ing,Longqing,H uaian red rice,Guan yun red rice
阴性
Negative
图47个水稻材料的酚反应结果
Fig. 4.Phenol r eaction of seven rice materials.
1―扬稻6号;2―日本晴;3―IR24;4―Dular;5―龙晴;6―02428;7-苏御糯。
1,Yangdao6;2,Nipponb are;3,IR24;4,Dular;5, Longqing;6,02428;7,Suyu nuo.
择。因此,开发基因功能标记对于提高水稻育种效率具有十分重要的意义。
多酚氧化酶是一类广泛分布于植物体内能催化多酚类氧化成醌类的金属酶。酶促褐变是农产品中普遍存在的一种有害现象,不仅影响产品的外观、风味、营养和加工性能,而且大大降低产品耐储性,易造成较大的经济损失[13]。籼稻栽培品种和一些野生稻的籽粒在苯酚溶液中变成棕色,这一过程类似于小麦面粉长期储藏褐变,因此,酚反应可能与水稻稻谷耐储藏性有关。目前,水稻多酚氧化酶基因已经克隆,已鉴定出4个酚反应阴性的等位基因,在水稻品种资源中主要是第3外显子缺失的两类等位基因[9-10]。
本研究根据多酚氧化酶等位基因的功能序列(functio n m otif)设计了功能标记FM ppo-18和FMppo-29。标记分析结果表明,只要PP O基因缺失18对碱基或缺失29对碱基,其酚反应表现型均为不染色,而携带正常PP O基因的材料酚反应均为染色。缺失带型不染色,非缺失带型染色,标记基因型与酚反应表现型完全对应。说明可以利用这些标记进行水稻酚反应相关性状的分子标记辅助选择。
本研究发现来源于国际水稻研究所的籼稻品种IR24,酚反应与其他材料不一样,处理48h后,稃尖和两片护颖先着色,而颖壳几乎不染色,处理72h 后,颖壳染色。IR24的酚反应可能还存在其他的调控因子,有待进一步研究证实。
3.2杂草稻与栽培稻的基因漂移
杂草稻(Ory z a sativ a f.sp ontanea)是具有杂草特性的水稻,其生育繁殖已脱离人类劳作控制范围,常与栽培稻伴生,在中国和世界大多数水稻生产国家和地区普遍发生,特别是南亚、东南亚,美国南部、北部,欧洲南部[17]。杂草稻在分类学上与栽培稻同属一个种(O.sativ a),但其主要特征是具有强落粒性和休眠性,这些特征为其广泛分布提供了可能。杂草稻作为一种水稻田间杂草,使得水稻减产,影响稻米品质。绝大多数杂草稻的果皮为红色,所以又被称为红米稻。杂草稻形态上变异较大,介于栽培稻与野生稻之间。长期共生于同一生态区域,栽培稻与杂草稻通过天然异交和基因渐渗,两者存在许多相似性,使得相对于其他杂草更难以控制。因此,探索江淮流域杂草稻的起源进化对于杂草稻的控制具有重要意义。
41
杨杰等:水稻多酚氧化酶基因功能标记的开发与应用
穞稻、塘稻等20世纪50年代江淮流域发现的杂草稻以及普通野生稻,稻壳都是黑色,酚反应也是黑色。两个标记分析结果为非缺失带型,表明他们携带正常功能的P PO基因。近年来危害江苏省水稻生产的16份红米杂草稻材料,稻壳为草黄色,其中14份材料酚反应阳性,2份酚反应阴性,表现18 bp缺失带型。本研究发现太湖流域粳稻尤其是目前粳稻生产的主要品种是P PO基因发生18bp的缺失,2份来自江苏淮安、连云港灌云的杂草稻携带18bp缺失的PP O等位基因,推测其可能与粳稻存在基因交流。我们利用卢宝荣等[19]开发的可以鉴别籼粳特征的34对InDel标记进行分析,结果表明多数近年来收集的杂草稻的籼稻指数为0.7左右(未发表结果),说明这些杂草稻偏籼型,但也有粳稻成分。我们发现近年来危害江苏省水稻生产的杂草稻柱头外露率高,粳稻花粉向杂草稻漂移是可能的;同时这两份杂草稻主要来自江苏省的苏北地区,这一地区是中熟中粳种植区,其抽穗期与杂草稻接近,所以通过天然异交发生基因漂移是完全可能的。3.3酚反应与水稻分类
Oka等[18]的研究认为酚反应是水稻籼粳分类的重要依据之一,一般籼稻酚反应为阳性,即稻壳可被染色,粳稻酚反应为阴性,稻壳不染色。本研究发现19份栽培稻中除Dular和龙晴这2份材料外,其他材料的酚反应结果与籼粳分类的结果是一致的。Dular是来自印度Aus群的材料,具有广亲和基因S5-n[20],传统分类认为是籼稻。本研究发现Dular 携带29bp缺失的多酚氧化酶基因,酚反应呈阴性,说明Dular有粳稻血缘。利用卢宝荣等[19]开发的34对InDel标记鉴定表明其籼稻指数为0.74,表明Dular有26%的粳稻血缘,与传统认为Dular属籼粳中间型材料的观点相吻合。龙晴是籼粳交后代,株叶形态偏籼型,酚反应呈阴性,我们的研究发现该材料的细胞质为粳稻来源[21],因此,龙晴应是籼粳中间型材料。江淮流域早年发现的杂草稻即穞稻和塘稻分类上属粳稻,细胞质也与粳稻相同,而近年来发现的红米杂草稻细胞质与籼稻相同,但这23份江淮流域杂草稻中21份材料的酚反应为阳性,与3份普通野生稻的基因型和酚反应表现型一致。多酚氧化酶基因可能与水稻等植物抗病虫以及储藏等特性有关,多数杂草稻都在自然选择中保留了功能正常的PP O基因,可能与杂草稻的野生适应特性有关。
参考文献:
[1]Kato S,K osak a H,H ara T.On th e affinity of rice varieties as
sh own by fertility of hybrid plants.B ull S ci Fac A g ric K y u-shu Univ,1928,3:132-147.
[2]Oka H I.Intervarietal variation and classification of cultivated
rice.Ind ian J Genet P lant B reed,1958,18:79-89.
[3]Oka H I.Phylogen etic differentiation of the cultivated rice
plant:?.Variations in respective characteristics and their com binations in rice cultivars.Jp n J B reed,1953,3:33-43.
[4]Morish ima H,Oka H I.Th e pattern of interspecific variation
in the genus Ory z a:Its q u antitative r epresen tation by statist-i cal meth ods.E volution,1960,14:153-165.
[5]McCouch S R,Kochert G,Yu Z H,et al.Molecular mapping
of rice ch rom osomes.T heor A p p l Genet,1988,76:815-829.
[6]T anksley S D,Fulton T M,McCouch S R.Genetic M aps:
Locu s M aps of Comp lex Genom es.Cold Spr ing H arbor,NY: Cold Sprin g H arbor Laboratory Press,1993.
[7]Lin S Y,Nagamura Y,Ku rata N,et al.DNA mark ers tigh tly
lin ked to genes,P h,A lk and R c.R ice G enet N ew sl,1994, 11:108-109.
[8]S aito A,Yano M,Kish imoto N,et al.Link age map of restric-
tion fragmen t length polymorphism loci in rice.J p n J B reed, 1991,41:665-670.
[9]Yu Y H,T ang T,Qian Q,et al.Indep endent losses of func-
tion in a polyph enol oxidase in rice:Differentiation in grain discoloration betw een sub species and th e role of positive selec-tion under domestication.P la nt Cell,2008,20:2946-2959.
[10]Briana L G,Karl J S,Kenneth M O.Novel P hr1mutations
and th e evolu tion of phen ol reaction variation in US w eedy rice (Oryz a sativa).N ew P hytol,2009,184(4):842-850. [11]T hipyapong P,Hu nt M D,S teffens J C.Systemic w ound in-
duction of potato(S olanum tuberosum)polyph enol oxidase.
P hytochemistr y,1995,40:673-676.
[12]Li L,Steffens J C.Overexp ression of polyph enol ox idase in
transgenic tom ato plants results in en han ced bacterial disease resistance.P lanta,2002,215:239-247.
[13]王曼玲,胡中立,周明全,等.植物多酚氧化酶的研究进展.
植物学通报,2005,22(2):215-222.
[14]Andersen J R,L bb erstedt T.Functional markers in plan ts.
T ren d s P lant S ci,2003,8(11):554-560.
[15]王丰,李金华,柳武革,等.一种水稻香味基因功能标记的
开发.中国水稻科学,2008,22(4):347-352.
[16]陆艳婷,刘庆龙,王俊敏,等.利用等位基因特异扩增快速检
测水稻香味基因.作物学报,2008,34(2):243-246.
[17]蒋菏,吴竞仑,王根来.连云港穞稻研究.作物品种资源,
1985(2):4-7
[18]Oka H I,Chang W T.Rice varieties intermed iate betw een
w ild an d cultivated forms and the origin of the japonica type.
B ot Bull A cad Sin(T aiwan),1962,3:109-131.
[19]卢宝荣,蔡星星,金鑫.籼稻和粳稻的高效分子鉴定方法及
其在水稻育种和进化研究中的意义.自然科学进展,2009,19
(6):628-638.
[20]Ch en J J,Ding J H,Ouyang Y D,et al.A triallelic system of
S5is a major regulator of the reproductive barrier an d compat-i bility of indica-japonica hybrids in rice.P roc N atl A cad S ci US A,2008,105(32):11436-11441.
[21]杨杰,王军,曹卿,等.江淮流域杂草稻叶绿体DNA
的籼粳分化.中国水稻科学,2009,23(4):391-397.
42中国水稻科学(Chin J Rice Sci)第25卷第1期(2011年1月)
麻醉药物基因组学进展
麻醉药物基因组学进展 本文对药物基因组学的基本概念和常用麻醉药的药物基因组学研究进 展实行综述。 药物基因组学是伴随人类基因组学研究的迅猛发展而开辟的药物遗传 学研究的新领域,主要阐明药物代谢、药物转运和药物靶分子的基因 多态性及药物作用包括疗效和毒副作用之间关系的学科。 基因多态性是药物基因组学的研究基础。药物效应基因所编码的酶、 受体、离子通道作为药物作用的靶,是药物基因组学研究的关键所在。基因多态性可通过药物代谢动力学和药物效应动力学改变来影响麻醉 药物的作用。 基因多态性对药代动力学的影响主要是通过相对应编码的药物代谢酶 及药物转运蛋白等的改变而影响药物的吸收、分布、转运、代谢和生 物转化等方面。与麻醉药物代谢相关的酶有很多,其中对细胞色素- P450家族与丁酰胆碱酯酶的研究较多。基因多态性对药效动力学的影 响主要是受体蛋白编码基因的多态性使个体对药物敏感性发生差异。 苯二氮卓类药与基因多态性:咪唑安定由CYP3A代谢,不同个体对咪 唑安定的清除率可有五倍的差异。地西泮是由CYP2C19和CYP2D6代谢,基因的差异在临床上可表现为用药后镇静时间的延长。 吸入麻醉药与基因多态性:RYR1基因变异与MH密切相关,现在已知 至少有23种不同的RYR1基因多态性与MH相关。氟烷性肝炎可能源于 机体对在CYP2E1作用下产生的氟烷代谢产物的一种免疫反应。 神经肌肉阻滞药与基因多态性:丁酰胆碱酯酶是水解琥珀酰胆碱和美 维库铵的酶,已发现该酶超过40种的基因多态性,其中最常见的是被 称为非典型的(A)变异体,与用药后长时间窒息相关。 镇痛药物与基因多态性:μ-阿片受体是阿片类药的主要作用部位, 常见的基因多态性是A118G和G2172T。可待因和曲马多通过CYP2D6代
浅谈我国转基因水稻的研究(一)
浅谈我国转基因水稻的研究(一) 论文关键词]水稻转基因论文摘要]稻转基因研究是国内外植物分子遗传学研究的热点之一。目前,水稻转基因研究在我国已取得显著进展。详细介绍转基因技术,并阐明我国转基因技术在水稻上的应用及研究进展, 水稻是我国的重要经济作物和粮食作物。水稻分布极其广泛,由于生态环境的复杂性和所处地理环境的影响,水稻在漫长的进化过程中,形成了极其丰富的遗传多样性,染色体组型和数目复杂多样,成为研究稻种起源、演化和分化必不可少的材料。 植物转基因技术是利用遗传工程手段有目的地将外源基因或DNA构建,并导入植物基因组中,通过外源基因的直接表达,或者通过对内源基因表达的调控,甚至通过直接调控植物相关生物如病毒的表达,使植物获得新性状的一种品种改良技术。它是基因工程、细胞工程与育种技术的有机结合而产生的一种全新的育种技术体系。转基因技术可以将水稻基因库中不具备的各种抗性或抗性相关基因转入水稻,进一步拓宽了水稻抗病基因源,为抗病育种提供了一条新途径。 一、国内外的转基因技术 转基因技术自20世纪70年代诞生以来,已经取得迅速的发展。到目前为止,中国已经是全球第4大转基因技术应用国。 转基因生物技术的应用,大多分布在抗虫基因工程、抗病基因工程、抗逆基因工程、品质基因工程、品质改良基因工程、控制发育的基因工程等领域。中国是继美国之后育成转基因抗虫棉的第二个国家。现在河北省与美国孟山都合作育成33B抗虫棉(高抗棉铃虫、抗枯萎病、耐黄萎病)。由中国农科院生物中心、江苏省农科院导入Bt基因,由安徽省种子公司,安徽省东至县棉种场共同选育的抗虫棉“国抗1号”在安徽省已通过审定。国际水稻所将抗虫基因导入水稻,育成抗二化螟、纵卷叶螟的转基因水稻。中国农科院、中国农业大学、中国科学院、河南农科院等许多科研单位和高校将几丁质酶和葡聚糖酶双价基因导入小麦育成抗病转基因小麦、转基因烟草、转基因水稻等等。英国爱丁堡大学将水母发光基因导入烟草、芹菜、马铃薯等作物,获得发光作物,驱赶害虫。 至于油菜方面利用转基因工程培育雄性不育系及其恢复系的研究,亦取得了突破性的进展。比利时为了提高菜饼粗蛋白质的含量,将一种草控制的蛋白质基因转移到油菜上来,选出高蛋白质含量的转基因油菜品种。瑞典Svalow-Weibull等公司利用基因工程技术将外源基因导入甘蓝型油菜,培育成抗除草剂油菜新品种;比利时PGS公司采用基因工程手段创造出新的油菜授粉系统;法国应用原生质体融合技术将萝卜不育细胞质的恢复基因引入甘蓝型油菜,充分利用萝卜不育细胞质不育彻底的特性,实现了萝卜不育细胞质的三系配套,对推动全球杂交油菜育种具有革命性的影响。 二、我国转基因技术在水稻上的应用及研究进展 我国是农业超级国,因此,中国人吃饭问题的关键是水稻问题(高产和抗性问题),而水稻问题的核心便是转基因技术在水稻中的成功应用。 近年来,植物抗病毒基因工程的技术路线已趋向成熟,国内外相继开展了水稻东格鲁病、条纹叶枯病、黄矮病、矮缩病等8种病毒病的转基因育种研究,将各病原病毒的外壳蛋白基因、复制酶基因、编码结构或非结构蛋白基因干扰素CDNA等分别导入水稻,获得了抗不同病毒病的转基因株系或植株。在我国,转基因技术在水稻中的应用已经取得了惊人的成果。(一)转基因技术在提高水稻植株的抗Basra除草剂的成果 王才林等利用花粉管通道法将抗Basta除草剂的bar基因导入水稻品系“E32”,获得转基因植株。抗性鉴定表明,转基因植株能充分表达对Basta除草剂的抗性;通过对转基因植株后代PCR分析,证实bar基因已整合到受体植株的基因组中,遗传分析表明,bar基因能在有性生殖过程中传递给后代,并在T代开始分离出抗性一致的稳定株系。段俊等利用转基因技术,
转基因作物的研究进展
生物与环境工程学院课程论文 转基因作物的研究进展 学生姓名:魏斌聪 学号:200806016139 专业/班级:生物工程081班 课程名称:生物工程原理 指导教师:陈蔚青教授 浙江树人大学生物与环境工程学院 2011年5月
转基因作物的研究进展 魏斌聪 (浙江树人大学生物与环境工程学院生工081班浙江杭州310015) 摘要:人们将所需要的外源基因(如高产、抗病虫害优质基因) 定向导入作物细胞中, 使其在新的作物中稳定遗传和表现,产生转基因作物新品种, 是大幅度提高作物产量的一项新技术。本文先描述了转基因作物的发展进程,对其基因问题的研究作了讨论,并列出转基因作物目前存在的主要问题并作分析,最后对此项技术作出展望。 关键词:转基因作物;DNA技术;基因导入;安全性 前言 转基因植物(transgenic plant),是指基因工程中运用DNA 技术将外源基因整合于受体植物基因组、改变其遗传组成后产生的植物及其后代。转基因植物的研究主要在于改进植物的品质,改变生长周期等提高其经济价值或实用价值。[ 1 ]其主要范围是在作物方面,如可食用的大豆、玉米等,或者可投入生产的棉花等作物。 从表面上看来,转基因作物同普通植物似乎没有任何区别,它只是多了能使它产生额外特性的基因。从1983年以来,生物学家已经知道怎样将外来基因移植到某种植物的脱氧核糖核酸中去,以便使它具有某种新的特性:抗除莠剂的特性,抗植物病毒的特性,抗某种害虫的特性。[ 2 ]这个基因可以来自于任何一种生命体:细菌、病毒、昆虫等。这样,通过生物工程技术,人们可以给某种作物注入一种靠杂交方式根本无法获得的特性,这是人类9000年作物栽培史上的一场空前革命。[ 3 ] 1 转基因作物的发展进程 转基因作物的研究最早始于20世纪80年代初期。1983年,全球第一例转基因烟草在美国问世。1986年,首批转基因抗虫和抗除草剂棉花进入田间试验。1996年,美国最早开始商业化生产和销售转基因作物(包括大豆、玉米、油菜、
转基因育种研究进展
作物转基因育种研究进展 摘要:近年来,植物基因工程取得了辉煌的成就,而转基因技术由于其巨大的产业价值,特别是在作物品质改良、产量和抗逆性提高等方面的明显优势,一直是国际农业高新技术竞争的焦点和热点。本文主以棉花、玉米、水稻为例就转基因育种技术在作物上的研究进展进行相关的介绍。 关键词:作物,棉花,玉米,水稻,转基因育种,研究进展 植物转基因技术是指利用重组技术、细胞DNA培养技术或种质系统转化技术将目的基因导入植物基因组,并能在后代中稳定遗传,同时赋予植物新的农艺性状,如抗虫、抗病、抗逆、高产、优质等。常规育种常常受有性杂交亲和性的制约,而利用转基因技术可以打破物种界限、克服有性杂交障碍,快速有效地创造遗传变异,培育新品种、创造新类型,大大缩短新品种育成的时间。因此,随着现代生物技术的迅速发展,植物转基因技术也蓬勃发展[1]。 1 转基因棉花育种的研究与进展 近年来,随着基因工程技术的不断发展,利用生物技术来创新棉花种质资源和培育新品种是一条非常有效的途径,极大地推动了棉花遗传育种的发展[2]。中棉所是世界上唯一可以同时采用农杆菌介导法、花粉管通道法、基因枪轰击法快速获得转基因抗虫棉新材料的技术平台,能将植物嫁接技术成功应用于转基因棉花的快速移栽,成活率超过90%。未来3~5年,中棉所将挖掘、整合与优化抗病、抗除草剂等基因10个,筛选高产因子、高品质纤维等基因或分子标记150个,创造转基因棉花育种新材料100份以上,培育重大新品种(组合)3~5个。 1.1转抗虫基因 1991年成功将外源Bt基因导人棉株中,1992年人工合成了全长1824bp的CrylAb和CrylAc融合的GFMCry1A基因,并于1993年采用农杆菌介导法和外源基因胚珠直接注射法成功导入晋棉7号、中棉12、泗棉3号等主栽品种,获得了高抗棉铃虫的转基因棉花株系;包含CryIAc和AP基因双价抗虫基因载体,通过农杆菌介导转化冀合321胚性愈伤组织,经6代筛选后培育出抗棉铃虫90%的纯合品系,且农艺性状均优于对照。 1.2转抗黄萎病相关基因 利用花粉管通道法和农杆菌介导转化法将菜豆中的几丁质酶和烟草中的葡聚糖酶基因转入棉花,并从转基因高世代材料中筛选出了高抗黄萎病的品系;将天麻抗真菌蛋白基因用花粉管通道法转化天然彩色棉主栽品种,从高世代系中选育出既抗枯萎病又抗黄萎病的兼抗材料;将葡萄糖氧化酶基因(GO)转入棉花,转基因后代对枯萎病和黄萎病抗性均有显著提高,部分材料抗性达到抗病水平。1.3转抗除草剂基因 1997年由美国孟山都公司推出抗除草剂棉花抗性品种,他们从土壤农杆菌变种CP4中分离到编码抗草甘膦酶的基因,并通过农杆菌介导法转化珂字棉312,把该基因导入棉花植株,从而使其对草甘膦产生抗性。采用中棉35下胚轴为材料,将草甘膦突变基因aroAM12导入到棉花中,获得65棵再生植株,通过Southern及Western试验验证了该基因的导入和表达状况,结果表明,转化株对草甘膦具有很高的抗性;将抗草甘膦基因aroAM12和抗虫基因Btslm一起整合到一个载体中,并以抗草甘膦基因作为选择标记,通过转化棉花品种石远321后获得了抗草甘膦和抗棉铃虫的再生株。
氯吡格雷药物基因组学及个体化治疗研究进展与展望
·944· 中华老年多器官疾病杂志 2013年12月28日 第12卷 第12期 Chin J Mult Organ Dis Elderly, Vol.12, No.12, Dec 28, 2013 收稿日期: 2013?06?18; 修回日期: 2013?07?18 基金项目: 国家自然科学基金面上项目(30971259,30570736/C03030201); 解放军总医院临床扶持基金(2012FC-TSYS-3042) 通信作者: 卢才义, E-mail: cylu2000@https://www.wendangku.net/doc/081371257.html,; 尹 彤, E-mail: yintong2000@https://www.wendangku.net/doc/081371257.html, ·综 述· 氯吡格雷药物基因组学及个体化治疗研究进展与展望 张蓝宁,卢才义*,尹 彤* (解放军总医院老年心血管病研究所,北京 100853) 【摘 要】通过与阿司匹林联合应用,氯吡格雷已经成为治疗急性冠脉综合征和预防经皮冠状动脉介入术后支架内 血栓形成和再发缺血事件的经典口服抗血小板药物。尽管如此,氯吡格雷抗血小板的反应性和疗效存在显著的个体间差异。近年来的研究证实,除临床环境因素外,遗传变异是导致氯吡格雷抗血小板反应性个体间差异的重要因素之一。多项大规模临床药物基因组学研究发现,参与氯吡格雷代谢的关键酶——CYP2C19功能缺失型等位基因与氯吡格雷治疗期间高血小板反应性及心血管一级缺血终点事件的发生密切相关。另外,与氯吡格雷代谢相关的其他基因变异型也被证实可能与氯吡格雷抗血小板反应性及不良心血管事件相关。在此基础上,利用药物基因组学基因型检测指导氯吡格雷个体化抗血小板治疗,可能部分克服氯吡格雷治疗期间的高血小板反应性,但研究结果之间仍存在争议,尚需深入研究以提供更有力的证据。除此之外,未来有必要进一步深入研究基因型检测联合血小板功能监测共同指导氯吡格雷抗血小板个体化治疗的效果。 【关键词】氯吡格雷;遗传药理学;CYP2C19;血小板反应性;心血管缺血事件;个体化医学 【中图分类号】 R541.4 【文献标识码】 A 【DOI 】 10.3724/SP.J.1264.2013.00239 Pharmacogenomics and individualized therapy of clopidogrel: evidence and perspectives ZHANG Lan-Ning, LU Cai-Yi *, YIN Tong * (Institute of Geriatric Cardiology, Chinese PLA General Hospital, Beijing 100853, China) 【Abstract 】 Dual antiplatelet therapy with aspirin and clopidogrel is the standard care to prevent stent thrombosis and recurrent ischemic events after acute coronary syndrome or stent placement. However, there is a large inter-individual variability in biological anti-platelet responsiveness and clinical outcomes in patients after clopidogrel treatment. Apart from clinical and environmental factors, recently accumulated evidence strongly confirms the pivotal role of genetic factors for the variability of clopidogrel responsiveness. Several large-scale pharmacogenomic studies found that the loss-of-function alleles of CYP2C19 and the key enzyme in clopidogrel metabolism are the predominant genetic mediators of low clopidogrel responsiveness and recurrent cardiovascular events. Other genetic polymorphisms related with clopidogrel metabolism may also contribute to the variability of clopidogrel efficacy. On the basis of these observations, it is still in controversy whether CYP2C19-genotype-guided individualized clopidogrel therapy could overcome the high on-treatment platelet reactivity to clopidogrel. In the future, it is necessary to combine genotyping and platelet function testing to guide the individualized clopidogrel therapy. 【Key words 】 clopidogrel; pharmacogenetics; CYP2C19; platelet function; cardiovascular ischemic events; individualized medicine This work was supported by the General Program of National Natural Science Foundation of China (30971259, 30570736/C03030201) and the Supporting Fund of People’s Liberation Army General Hospital (2012FC-TSYS-3042). Corresponding author: LU Cai-Yi, E-mail: cylu2000@https://www.wendangku.net/doc/081371257.html,; YIN Tong, E-mail: yintong2000@https://www.wendangku.net/doc/081371257.html, 通过与阿司匹林联合应用,氯吡格雷(clopidogrel )已经成为治疗急性冠脉综合征(acute coronary syndrome ,ACS )和预防经皮冠状动脉介入(percutaneous coronary intervention , PCI )术后支架内血栓形成和再发缺血事件的经典口服抗血小板药物[1,2], 但氯吡格雷抗血小板反应性和疗效存在显著的个体差异。除临床环境因素外,基因多态性在其中起了重要作用。多项大
转基因作物安全评价研究进展
转基因作物安全评价研究进展 转基因技术是现代生物技术的核心。推进转基因技 术研究与应用,是着眼于未来国际竞争和产业分工的重大发展战略,是解决粮食短缺、人口问题、确保国家粮食安全的必然要求和重要途径。温家宝总理2010年政府工作报告中 明确指出要重点抓好“以良种培育为重点,加快农业科技创新和推广,实施好转基因生物新品种培育科技重大专项”工作。“农业转基因生物新品种培育科技重大专项”的实施,标志着转基因技术已成为我国抢占科技制高点和增强农业国际竞争力的战略重点。转基因技术自诞生以来,生物安全问题相伴而生。在转基因作物的研究和产业化过程中,转基因作物的安全性成为亟待解决的关键问题。 1 国内外转基因作物安全评价原则 全球各国都加强了对转基因作物安全性评价的研究工作,主要国际组织和研究机构都制定了相关“基于实质等同性”的安全评价原则和标准,在遵循这一原则的基础上对转基因作物进行安全性评价…。 2转基因作物安全评价体外实验研究现状 目前,转基因作物食用安全性评价主要方法是实验研究法。实验研究法有体外实验和体内实验两种研究途径。体外实验是通过各种物理化学方法对转基因作物及其产品进行评价分析。主要有关键成分分析和营养学评价:如蛋白质及氨基酸、脂肪及脂肪酸、碳水
化合物、矿物质、维生素等营养成分分析;抗营养因子和酶抑制剂等抗营养成分和天然毒素分析;因基因修饰生成的新成分和其他可能产生的非预期成分分析等。还有转基因作物主要成分稳定性分析:如 加工贮存过程中转基因作物稳定性的研究;转基因作物在动物体内消化稳定性的研究等。 现有研究表明转基因大豆、豆粕中干物质、粗脂肪、粗蛋白、中性洗涤纤维、酸性洗涤纤维、灰分、钙和总磷8种普通营养成分与普通大豆含量较接近,无显著差异;转基因大豆中氨基酸、微量元素铁、铜、锰、锌含量与普通大豆相近。转基因大豆中转基因植酸磷、胰蛋白酶抑制因子、脲酶活性和蛋白溶解度等抗营养因子未发生变化,大豆异黄酮和大豆凝集素等在二者之间也具有实质等同性[10]。研究者 还认为尽管转基因大豆中转基因豆粕C14:1脂肪酸、C22:0 脂肪酸、共轭亚油酸含量存在差异,但二者差异没有实际意义,饱和脂肪酸、不饱和脂肪酸含量及各种脂肪酸含量与传统常规大豆间无显著差异。转基因大豆与常规大豆具有实质等同性。部分研究也表明转基因玉米、转基因大米与普通作物具有实质等同性。 3转基因作物安全评价体内实验研究现状 体内实验主要是通过先饲喂动物转基因产品,然后通过研究实验动物身体各方面机能参数(日常活动、体液指标、器官发育、病理检查等)来评价转基因作物的安全性。一些研究表明转基因作物对动物的影响与传统非转基因作物相同。如有研究证实:转基因大豆
药物基因组学相关数据库
药物基因组学数据库 1、Drugbank 2、dgidb 3、pharmGKB 4、cancercommon 5、ChEMBL 6、mycancergenome 7、TTD 8、guidetopharmcology 9、clearityfoundation 10、CIViC https://https://www.wendangku.net/doc/081371257.html,/#/home 11、DoCM https://www.wendangku.net/doc/081371257.html,/ 1 Drugbank 药物和药物靶标资源库。DrugBank是一个独特的生物信息学/化学信息学资源,它结合了详细的药物(例如化学制品)数据和综合的药物靶点(即:蛋白质)信息。该数据库包含了超过4100个药物条目,包括超过800个FDA认可的小分子和生物技术药物,以及超过3200个试验性药物。此外,超过1.4万条蛋白质或药物靶序列被链接到这些药物条目。每个DrugCard条目包含超过80个数据域,其中一半信息致力于药物/化学制品数据,另一半致力于药物靶点和蛋白质数据。许多数据域超链接到其他数据库(KEGG、PubChem、ChEBI、Swiss-Prot和GenBank)和各种结构查看小应用程序。该数据库是完全可搜索的,支持大量的文本、序列、化学结构和关系查询搜索。DrugBank的潜在应用包括模拟药物靶点发现、药物设计、药物对接或筛选、药物代谢预测、药物
相互作用预测和普通药学教育。DrugBank可以在http://www.drugbank.ca 使用。广泛应用于计算机辅助的药物靶标的发现、药物设计、药物分子对接或筛选、药物活性和作用预测等。 在查询中,每一种药物对应1个DrugCard,即我们所得到的检索结果。每一个DrugCard都包含的数据信息分为药物、靶标和酶三部分。 药物信息包括了该药物的CAS号、商品名、分子式、分子量、SMILES、2D 和3D结构、logP、logS、pKa、熔点、吸收性、Caco-2细胞穿透性、药物类别和临床使用、性质描述、剂型与给药途径、半衰期、体内的生物转化、毒性、作用于哪些生物体、食物对服用的影响、与其它药物的相互作用、作用机理、代谢途径、药理学特征、与蛋白质的结合情况、溶解度、物质形态、同义词、关于合成的相关文献等,还与ChEBI、GenBank、PubChem等外部数据库有链接。 靶标的信息包括ID、名称、靶标基因的名称、蛋白质序列、残基数目、分子量、等电点、功能和活性、参与的代谢途径和反应、体内分布、靶标信号、跨膜区域、靶标基因序列及其在GenBank、HGNC等外部数据库中的ID和链接、参考文献,以及在GenBank和Swiss-Prot中的链接。 酶的信息包括名称、蛋白质序列、基因名称、在Swiss-Prot 等数据库中的链接。 在DrugBank的主界面上,在Browse菜单下可以浏览数据库的内容,其中PharmaBrowse为用户提供了分类浏览的功能。这为药剂师、医生以及寻找潜在药物的研究人员提供了方便。在Search下拉菜单下,就是Drug Bank的4类检索方式。ChemQuery允许用户通过绘制结构图或书写SMILES、分子式进行结构搜索。在检索过程中还可以对搜索药物类型、分子量范围、搜索结果相似度、结果数量最大值等进行设置。TextQuery则为文本检索功能。文本检索支持逻辑运算符连接及在特定领域内搜索。例如,在“dextromethorphan”中检索混合物,可以键入“mixtures:dextromethorphan”,即用分号在后面输入领域,同时可以加入逻辑运算符,例如,在“dextrome thorphan”和“doxylamine”2个领域进行检索,可以键入“mixtures:dextromethorphan AND mixtures:doxylamine”。SeqSearch为用户提供了通过序列检索蛋白质的功能。Data Extractor是1
水稻转基因步骤
在植物转基因过程中,为了有效地识别和筛选转化子,常将目的基因和标记基因构建在同一表达载体中。这种载体结构导致转基因植物中目的基因和标记基因始终共存,而标记基因(尤其是抗生素抗性基因)的存在可能给转基因植物的生物安全带来隐患。目前已研发了多种方法剔除转基因植物中的标记基因,其中最常见的是共转化法(Komari 1996,McCormac 等2001)。共转化系统是采用2个质粒或1个含有两套T—DNA表达盒的表达载体共同转化植物,其中一套表达盒含有抗性选择标记基因,另一套表达盒含有目的基因,它们转化植物时可能整合到植物基因组的不同位置。转基因植株在减数分裂过程中,标记基因和目的基因发生分离,从而可在转基因后代中筛选到只含目的基因而不含选择标记基因的个体。共转化从根本上排除了转基因植物中的选择标记,是保证人畜和环境安全的重要措施,因此受到了广泛的重视。Zhou 等(2003)认为,用分别含一个T-DNA区的两个载体共转化的效率低于双T-DNA区表达载体的共转化效率。目前关于利用双T-DNA区表达载体,获得无选择标记转基因阳性株系的研究已有不少报道(唐俐等2006,张秀春等2006,于恒秀等2005)。花药培养与遗传转化技术相结合,可以快速获得纯合转基因植株(斯华敏等,1999,付亚萍等,2001),但是应用花药培养快速获得只含目的基因而无选择标记的转基因研究尚未见报告。 水稻是最主要的粮食作物,转基因水稻的安全显得尤为重要。本实验室通过农杆菌介导的水稻转化体系,将包含人乳铁蛋白(hLF)、高赖氨酸(SB401)、高甲硫氨酸(RZ10)基因的表达载体p13HSR成功转化脆茎稻,由于该表达载体采用双T-DNA结构,将检测出含选择标记潮霉素磷酸转移酶基因(hpt)和目的基因的转基因阳性T0植株按单株直接进行花药培养。在189株二倍体花培植株中检出23株有目的基因没有选择标记hpt的转基因纯合植株,得率为9.87%。RT-PCR检测结果显示外源基因已整合到转基因水稻基因组中并转录。本文首次发现插入的外源基因间存在交换事件,从而改变了花培群体中无选择标记而目的基因阳性的转基因纯系的获得率。同时还对农杆菌介导的同一载体上多个基因转化水稻后,会出现个别基因丢失的情况进行了讨论。 基因转化方法参照Hiei等(1994)的方法并加以修改。取开花后12-15 d左右的稻穗脱粒,表面灭菌后接种在NB培养基上,26℃暗培养诱导愈伤组织。约5-7d后取愈伤组织在相同条件下继代培养,用于共培养。农杆菌于含50mg/L卡那霉素(Kam)的YM平板上划线,28℃黑暗培养3d,用金属匙收集农杆菌菌体,将其悬浮于共培养CM液体培养基中,调整菌体浓度至OD600为0.3-0.5,加入AS(终浓度为100mΜ),即为共培养转化水稻用的农杆菌悬浮液。将继代培养4d后的愈伤组织浸于此菌液中,20min后取出并用无菌滤纸吸去多余菌液,随即转入铺有无菌滤纸的固体培养基上,于26℃下暗培养2~3d。共培养后的愈伤组织在含有50mg/l潮霉素的筛选培养基上,26℃暗培养14d,再转到新鲜配制的筛选培养基上继续筛选14d。然后选择生长旺盛的抗性愈伤组织转移到含有50mg/l潮霉素的分化培养基上,暗培养3天后转至15h/d 光照条件下培养,再生的小苗在1/2MS上生根壮苗两周左右。选择高约10cm、根系发达的小
水稻转基因育种研究进展 7
水稻转基因育种研究进展 王彩芬,安永平,韩国敏,张文银,马 静 (宁夏农林科学院农作物研究所,宁夏永宁 750105) 摘要:对水稻转基因技术在抗虫、抗病、抗逆及改良米质等方面的进展进行了综述。 关键词:水稻; 转基因育种; 进展 中图分类号:S511.035.3 文献标识码:A 文章编号:1002-204X(2005)06-0055-03 20世纪下半叶以来,由于分子生物学研究的巨大成就,使生物学成为自然科学的带头学科,它的理论和方法已渗透到生命科学的许多领域,为生命科学的研究带来新的思维方式和研究手段。基因工程技术在植物遗传育种上应用很广泛,并取得了显著成就。 水稻是最重要的粮食作物之一,世界上约有一半以上的人口以稻米为主食。据专家预测,到2025年在现有稻谷产量的基础上再增加60%才能满足需要(K hush,1995)。随着人口的增长和耕地面积的减少,世界尤其是我国将面临粮食问题的严峻挑战,培育优良品种是提高稻谷产量的主要途径。传统的育种技术已为培育水稻新品种做出了巨大贡献,并将在今后继续发挥主导作用,但由于品种资源的贫乏,单靠传统育种已很难有大的突破。基因工程技术为水稻分子标记辅助育种、水稻转基因育种提供了一条新途径。转基因技术可以将水稻基因库中不具备的抗病、抗虫、抗除草剂、抗旱、耐盐、改善品质、提高产量等基因转入水稻,从而实现水稻种质创新和为生产提供优良品种。自1988年以来,国内外已得到了许多水稻转基因植株,涉及到抗虫、抗病、抗除草剂、抗旱、耐盐、改良品质等重要农艺性状,有些已进入田间试验和应用阶段。 1 水稻转基因育种进展 植物转基因育种是利用遗传工程的手段,有目的地将外源基因或DNA构建导入植物基因组,通过外源基因的直接表达,或通过对内源基因表达的调控,甚至通过直接调控植物相关生物如病毒的表达,使植物获得新的性状的一种品种改良技术。在植物分子生物学研究的众多材料中,水稻不仅是世界重要粮食作物,而且由于其基因组较小、重复序列较少的优点而成为一种重要的分子遗传学研究的单子叶模式植物,基因组测序已完成。自1988年首次获得转基因水稻以来,水稻转基因技术已获得突飞猛进的发展,目前已成功获得籼稻、粳稻、爪哇稻的转基因植物。随着基因枪转化技术的建立和根癌农杆菌介导转化法的成功,水稻基因转化技术日益完善。而且转移目标基因已从报告基因或筛选标记基因进入改良水稻抗性和适应性,以及改善品质,提高产量等重要基因的利用。 1.1 抗虫转基因水稻育种 水稻是虫害最多的大田作物,稻螟虫和稻飞虱危害最为严重,水稻中抗虫资源贫乏,转基因技术为抗虫品种的培育提供了一条新途径。自从1989年实现苏云金杆菌(Bacillus thuringiensis,简称Bt)抗虫基因转化水稻并得到再生植株以来,转抗虫基因水稻的研究取得了很大进展。转抗虫基因水稻包括转Bt基因、转蛋白酶抑制基因和转凝集素基因。在转Bt基因的研究方面,中国农科院生物技术中心杨虹等(1989)将Bt基因导入水稻品种台北309、中花8号的原生质体并获得再生植株;Fujim oto等(1993)通过电激法将cry LAb 基因导入水稻,首次报道了转Bt基因水稻对二化螟和稻纵卷叶螟的抗性。项友斌等(1999)利用农杆菌介导实现了苏云金杆菌抗虫基因cryI A(b)和cryI A(c)在水稻中的转化;黄健秋等(2000)利用农杆菌介导获得转(Bt)基因秀水11和春江11植株;薛庆中等(2002)利用农杆菌介导获得转双价抗虫基因(cryI Ac和豇豆胰蛋白酶抑制基因C pTI)浙大19植株;朱常香等(2002)获得Bt和X a21共转化水稻(C48)植株。近几年转Bt基因研究越来越多,进展很快,在籼稻、香稻、爪哇稻、杂交稻、深水稻中获得成功,选育出克螟稻1号、2号、3号(舒庆尧等,1998)。转Bt基因水稻在我国已进入环境释放阶段,有望培育出应用于生产的抗虫品种。 在转蛋白酶抑制剂基因水稻研究方面,通过电激介导原生质体转化,Xu等(1996)把豇豆胰蛋白酶抑制剂基因C pT i转入粳稻品种台北309,转基因植株对大螟和二化螟2种水稻虫害都具有抗性;通过基因枪介导马铃薯蛋白酶抑制剂基因PinⅡ转化水稻,Duan等(1996)获得了Nipponbare、台南67和Pi4等3个粳稻品种的抗大化螟转基因株系;Lee等(1999)利用PEG介导法将大豆K units胰蛋白酶抑制剂(SK TI)的cDNA转入粳稻Nagdongbyeo的原生质体,再生转基因植株的后代抗褐飞虱。曾黎琼等(2004)利用农杆菌介导将马铃薯蛋白酶抑制剂基因(PinⅡ)导入玉优1号、HT-7中;孔维文等(2004)利用农杆菌介导将PT A和马铃薯高赖氨酸蛋白基因(S B401)同时转入超级杂交稻亲本材料1826中。在转凝集素基因水稻研究中,主要是转雪莲花凝集素(G NA)基因,采用基因枪法,英国John Innes Centre(Maqbool等,1999;Rao等,1998;Sudhakar等,1998)把G NA基因导入AS D16、M5、M7、M12、FX92D、Basmati370等籼稻品种中,得到200多株转基因植株,G NA在水稻中呈高水平的组成性表达(用Ubi启动子)或韧皮部专一性表达(用Rssl启动子),转基因植株抗褐飞虱。在我国,傅向东等(1997)用G NA基因枪转化水稻IR72、IR76、珍汕97和秀水11等品种,部分转基因植株子代对褐飞虱有一定抗性;T ang(唐克轩等,1999)通过基因枪介导实现了G NA 基因和X a21基因的共转化,得到了转基因植株。唐克轩等(2003)利用农杆菌介导将半夏凝集素基因(pta)导入粳稻鄂宛105、中花12和籼稻E优532中,获得7个转基因纯系。 1.2 抗病转基因水稻育种 抗病转基因水稻包括转抗病毒基因、抗真菌病害基因和抗细菌病害基因。抗病毒转基因已开展了8种病毒的转基因研究,包括水稻通枯罗病毒(rice tungro disease)、水稻齿叶矮缩病毒(rice ragged 收稿日期:2005-07-21 作者简介:王彩芬(1968-),女,副研究员,从事水稻花培育种研究。T el:0951-*******E-mail:caifen-68@https://www.wendangku.net/doc/081371257.html,
转基因水稻大规模生产重组人血清白蛋白
转基因水稻大规模生产重组人血清白蛋白 由武汉大学生命科学院教授、武汉禾元生物科技有限公司董事长杨代常领衔的研发团队从2006年开始进行植物源替代血浆来源的医药蛋白的 研究与开发,现已取得突破性进展并已跨入规模化生产的阶段,填补了国际上此项技术空白。相关论文于2011年10月31日在线发表于《美国 科学院院报》。该论文在线之际,受到国外Scientist ,Nature news, The Australian, Thomson Reuters, Fox News, Agence France Presse (AFP法新社)等美国、英国、俄罗斯、德国、巴西、印度各专业杂志及媒体的广泛关注和报道。 该研究表明由转基因水稻种子生产的重组人血清白蛋白(OsrHSA)在生理生化性质、物理结构,生物学功能、免疫原性与血浆来源的人血清白 蛋白一致;并建立了大规模生产重组人血清白蛋白的生产工艺,获得了高纯度和高产量重组人血清白蛋白产品。利用大量数据证明了转基因 水稻种子可取代现有基于发酵的表达技术来生产重组蛋白质是经济有效的。正如PNAS 审稿人对该文章的评价:“这篇文章解决了在科学上振 奋人心、在经济上都非常重要的议题--即用转基因植物生产血浆产品或其他蛋白产品的技术平台,可代替其他基于发酵的表达技术,其重 要性也不言而喻……这篇文章近乎完美地证实了植物生产的医药蛋白和批准临床使用的血浆来源医药蛋白是完全相同的,并提供了翔实数据 证明植物系统规模化容易和成本优势。” 目前,人血清白蛋白(human serum albumin)广泛应用于临床治疗和细胞培养领域。常见的人血清白蛋白大多数从人的血浆中提取,这样的生 产方式不仅受到血浆供应的限制,而且还具有携带病毒传播的高风险性。国际上以重组人血白蛋白替代血源产品的应用已成为趋势,国内市 场需求也逐年扩大,2010年已达150吨。尽管市场广阔,但高纯度重组人血白蛋白的规模化生产技术和质量控制技术却是世界性难题。武汉禾 元历经多年的技术攻关,利用水稻胚乳表达技术平台,研发出国际先进水平的重组人血白蛋白产品生产技术,并成功实现重组人血白蛋白规 模化和产业化,完全摆脱了相关制约,具有纯度更高、无动物组分、安全、高效、绿色环保、廉价、无限量供应等优势。随着植物源重组人 血清白蛋白的发展,我国人血清白蛋白日益紧张的局面必将得到缓解。
基因芯片技术及其应用简介(精)
基因芯片技术及其应用简介 生物科学学院杨汝琪 摘要:随着基因芯片技术的发展,基因芯片越来越多的被人们利用,它可应用于生活中的方方面面,如:它可以应用于医学、环境科学、微生物学和农业等多个方面,基因技术的发展将有利于社会进一步的发展。 关键词:基因芯片;技术;应用 基因(gene是载有生物体遗传信息的基本单位,存在于细胞的染色体(chromosome上。将大量的基因片段有序地、高密度地排列在玻璃片或纤维膜等载体上,称之为基因芯片(又称DNA 芯片、生物芯片。在一块1 平方厘米大小的基因芯片上,根据需要可固定数以千计甚至万计的基因片段,以此形成一个密集的基因方阵,实现对千万个基因的同步检测。基因芯片技术是近年来兴起的生物高新技术,把数以万计的基因片段以显微点阵的方式排列在固体介质表面,可以实现基因检测的快速、高通量、敏感和高效率检测,将可能为临床疾病诊断和健康监测等领域,带来全新的技术并开拓广阔的市场。 1 基因芯片技术原理及其分类 1.1基因芯片的原理: 基因芯片属于生物芯片的一种"其工作原理是:经过标记的待测样本通过与芯片上特定位置的探针杂交,可根据碱基互补配对的原则确定靶序列[1],经激光共聚集显微镜扫描,以计算机系统对荧光信号进行比较和检测,并迅速得出所需的信息"基因芯片技术比常规方法效率高几十到几千倍,可在一次试验中间平行分析成千上万个基因,是一种进行序列分析及基因表达信息分析的强有力工具。 1.2基因芯片分类: 1.2.1根据其制造方法可分原位合成法和合成后点样法;
1.2.2根据所用载体材料不同分为玻璃芯片!硅芯片等; 1.2.3根据载体上所固定的种类可分为和寡核苷酸芯片两种; 1.2.4根据其用途可分测序芯片!表达谱芯片!诊断芯片等 2 基因芯片技术常规流程 2.1 芯片设计根据需要解决的问题设计拟采用的芯片,包括探针种类、点阵数目、片基种类等。 2.2 芯片制备将DNA, cDNA或寡核昔酸探针固定在片基上的过程。从本质上可分为两大类fz} ,一类是在片基上直接原位合成,有光蚀刻法、压电印刷法和分子印章多次压印法三种;另一类是将预先合成的探针固定于片基表面即合成点样法。 2.3 样品制备常规方法提取样品总RNA,质检控制。再逆转录为。DNAo 2.4 样品标记在逆转录过程中标记荧光素等。 2.5 芯片杂交标记的cDNA溶于杂交液中,与芯片杂交。 2.6 芯片扫描一用激光扫描仪扫描芯片。 2.7 图像采集和数据分析专用软件分析芯片图像,然后对数据进行归一化,最后以差异为两倍的标准来确定差异表达基因。 2.8 验证用定量PCR或原位杂交验证芯片结果的可信性。 3基因芯片合成的主要方法 目前已有多种方法可以将基因片段(寡核苷酸或短肽固定到固相支持物上。这些方法总体上有两种: 3.1原位合成:
麻醉领域的个体化用药,药物基因组学(Evan Kharasch)
Pharmacogenetics in Anesthesia Evan D. Kharasch, M.D., Ph.D. St. Louis, Missouri 302 Page 1 Pharmacogenetics (or pharmacogenomics) aims to understand the inherited basis for variability in drug response. The promise of pharmacogenetics has been a change from “one drug and dose fits all” to individualized predictive medicine, or “the right drug at the right dose in the right patient”. Anesthesiology as a specialty played a key role in developing pharmacogenetics. Prolonged apnea after succinylcholine, thiopental-induced acute porphyria, and malignant hyperthermia were clinical problems of the 1960’s whose investigation helped craft the new science of pharmacogenetics. Today we perhaps take for granted the knowledge that they are genetically-based problems, due to variants in pseudocholinesterase, heme synthesis and the ryanodine receptor, respectively. This review will address basic principles of pharmacogenetics and their application to drugs used in anesthetic practice. The term pharmacogenetics was originally defined (1959) as “the role of genetics in drug response”. Since the science of pharmacokinetics (drug absorption, distribution, metabolism, excretion) evolved earlier than pharmacodynamics, early pharmacogenetic studies addressed mainly pharmaco-kinetics. Application (fusion) of the genomic revolution and associated technologies to pharmaco-genetics spawned pharmacogenomics. Pharmacogenetics has been used by some in a more narrow sense, to refer only to genetic factors which influence drug kinetics and dynamics (drug receptor actions), while pharmacogenomics has been used more broadly to refer to the application of genomic technologies (whole-genome or individual gene changes) to drug discovery, pharmacokinetics and pharmacodynamics, pharmacologic response, and therapeutic outcome. Nonetheless, many consider this distinction unimportant and use the two terms interchangeably, as will this review. BASIC CONCEPTS A polymorphism is a discontinuous variation in a population (a bimodal or trimodal distribution). It is different than simple continuous variability (i.e. a unimodal population distribution, even if quite wide). A genetic polymorphism is the presence of multiple discrete states (i.e. for a particular trait) within a population, which has an inherited difference. The complete human genome consists of approximately 3 billion base pairs, which encode approximately 30,000 genes. A single nucleotide polymorphism (SNP) is a variation in the DNA sequence which occurs at a specific base. Polymorphisms are relatively common, occurring by definition in ≥1% of the population, while mutations are less common, occurring in <1%. Only 3% of DNA consists of sequences which code for protein (exons). Other portions of the DNA include promoter regions (near the transcription initiation site), enhancer regions (which bind regulatory transcription factors), and introns (DNA sequences which do not code for protein). After exons and introns are transcribed, the intronic mRNA is excised and the exonic mRNA is spliced together to form the final mature mRNA, which then undergoes translation into protein. SNPs are frequent, occurring in approximately 1:100-1:1000 bases. SNPs and mutations may occur in the coding or noncoding regions of the DNA. Since most occur in the latter, they are usually synonymous (or silent, having no effect on proteins), although intronic changes and promoter variants can change protein expression. Non-synonymous SNPs result in a change in an amino acid. A conservative change results in a similar amino acid that does not alter protein function, while a non-conservative change yields an amino acid which alters protein structure or function. These latter SNPs may be clinically significant. SNPs are not the only events which can cause RNA and protein changes; others are deletions, insertions, duplications, and splice variants, however these are not inherited. Multiple SNPs can occur in the DNA which encodes a particular protein. A haplotype is a set of closely linked alleles or DNA polymorphisms which are inherited together. While SNPs are important, haplotypes are more clinically relevant. Polymorphisms can be classified at the DNA locus (which depicts the normal “wild-type” and the altered base pair; for example the mu opioid receptor gene polymorphism at base pair 118 which codes for changing an adenine nucleotide to a guanine is abbreviated as A118G, or 118 A>G); at polymorphism changes the amino acid at position 40